Aim 2. Sensitivity of Australian bats to WNS mortality
Measure critical thermal and metabolic physiological traits
WNS can cause widespread physiological disruption in hibernating bats, thereby impact torpor arousal frequency (Fig. 1). How long a bat stays in torpor for depends on the environment, and ultimately predicts how long bats can be infected by Pd. Torpid and normothermic bats will be measured to obtain mean responses to temperature and humidity for the following key variables: resting metabolic rate, torpid metabolic rate, water loss rate; minimum body temperature in torpor, cooling rate during torpor entry, rewarming rate during arousal, and metabolic cost of interbout arousals. These data will allow us to calculate the energy balance of wild hibernating bats from observed patterns of torpor-arousal under different environmental conditions.
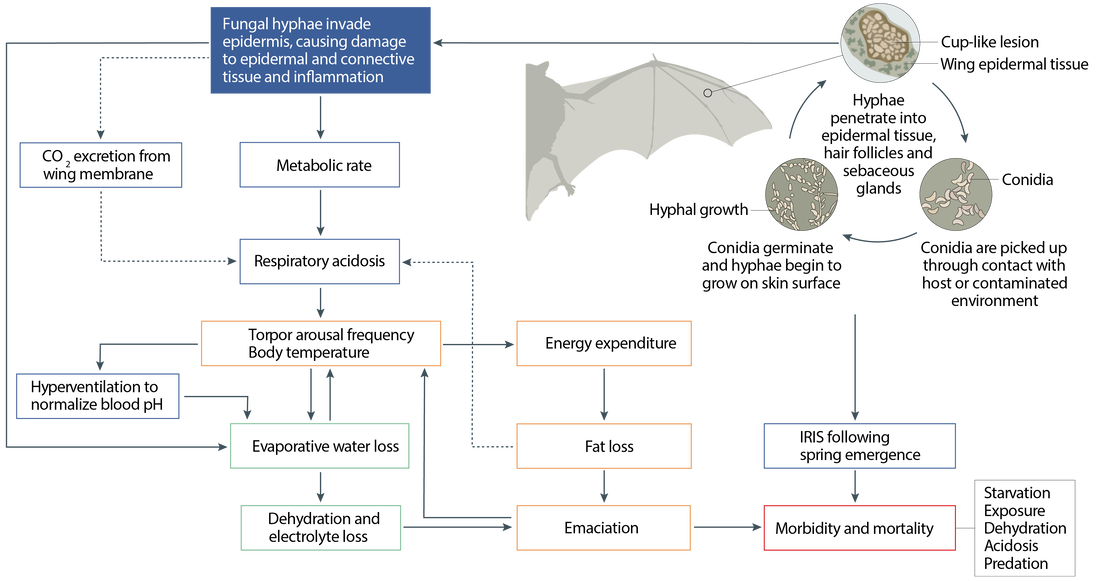
Fig. 1 | Effects of Pseudogymnoascus destructans infection on bat hosts. Image from Hoyt et al. (2021).
Roosting behaviour and winter activity
Cave-roosting insectivorous bats in southeastern Australia increase their body mass (fatten) in late Autumn and greatly reduce their activity during a period of hibernation over winter. However, little is known about their roosting behaviour or the frequency and the amount and purpose of activity over the winter season. Bat winter activity will be measured at key winter cave roosts covering different climates for each species using automated logging of ultrasonic bat calls.
- Acoustic detectors will be placed inside the cave to record aroused bats and immediately outside of winter roosts to record emerging bats over a sample of nights from autumn to spring.
- We will also use infrared illuminated time-lapse photography to record daily patterns of movement (indicating arousal) by hibernating bats.
Torpor-arousal cycles and winter energetics
Currently there is no published information on the thermoregulatory behaviour of any Australian cave-roosting bat. We will capture a sample of bats to measure body condition in late autumn, early hibernation, and late hibernation. We will use these data to quantify seasonal changes in body condition for each species. Extent of pre-hibernation fattening appears to correlate with geographic variation in winter severity and could provide a useful index of winter energy stress among bat populations. Sufficient body fat stores have been key to survival of WNS by impacted bat populations in North America.
We will measure individual torpor-arousal patterns of bats using temperature-sensitive radio telemetry and infrared time-lapse photography (as above). Miniature temperature-sensitive transmitters will be used to record continuous traces of skin temperature over winter in undisturbed hibernating bats. Temperature telemetry of a minimum sample of bats from populations inhabiting different climates will require a substantial amount of work, but these data are a priority because they provide detailed individual-based information on torpor-arousal patterns over weeks to months in relation to environmental conditions, which is needed to inform biophysical models about variation in hibernation energetics across species’ ranges.
We will measure individual torpor-arousal patterns of bats using temperature-sensitive radio telemetry and infrared time-lapse photography (as above). Miniature temperature-sensitive transmitters will be used to record continuous traces of skin temperature over winter in undisturbed hibernating bats. Temperature telemetry of a minimum sample of bats from populations inhabiting different climates will require a substantial amount of work, but these data are a priority because they provide detailed individual-based information on torpor-arousal patterns over weeks to months in relation to environmental conditions, which is needed to inform biophysical models about variation in hibernation energetics across species’ ranges.